Garry T. Cole
These fungi grow as saprophytes, parasites, or both by using specific proteolytic, glycolytic, or lipolytic enzymes to extracellularly break down substrates and to absorb the products of digestion through the fungal cell envelope.
The fungal cell wall gives shape and form, protects against mechanical injury, prevents osmotic lysis, and provides passive protection against the ingress of potentially harmful macromolecules.
Fungi are different from the Actinomycetes, a group of prokaryotic filamentous bacteria having peptidoglycans in their cell walls and an absence of nuclear membranes and organelles, but the two groups of microorganisms are usually considered together in texts.
Hyphal extension growth occurs apically by a sophisticated organization of tip-growth-related organelles and cytoskeletal elements. Hyphal wall and yeast cell wall polysaccharide synthetases are active at sites where growth is occurring and inactive when no growth is occurring. Morphogenesis is a balance between wall synthesis and wall lysis.
Sexual reproduction occurs by the fusion of two haploid nuclei (karyogamy), followed by meiotic division of the diploid nucleus. The union of two hyphal protoplasts (plasmogamy) may be followed immediately by karyogamy, or it may be separated in time.
Asexual reproduction occurs via division of nuclei by mitosis. With the absence of meiosis, other mechanisms associated with the nuclear cycle result in recombination of hereditary properties and genetic variation.
Macroscopic fungi such as morels, mushrooms, puffballs, and the cultivated agarics available in grocery stores represent only a small fraction of the diversity in the kingdom Fungi. The molds, for example, are a large group of microscopic fungi that include many of the economically important plant parasites, allergenic species, and opportunistic pathogens of humans and other animals. They are characterized by filamentous, vegetative cells called hyphae. A mass of hyphae forms the thallus (vegetative body) of the fungus, composed of mycelium. The more phylogenetically primitive molds (e.g., water molds, bread molds, and other sporangialsaclikeforms) produce cenocytic filaments (multinucleate cells without cross-walls), while the more advanced forms produce hyphae with cross-walls (septa) that subdivide the filament into uninucleate and multinucleate compartments. The septum, however, still provides for cytoplasmic communication, including intercellular migration of nuclei. Many fungi occur not as hyphae but as unicellular forms called yeasts, which reproduce vegetatively by budding. Some of the opportunistic fungal pathogens of humans are dimorphic, growing as a mycelium in nature and as a vegetatively reproducing yeast in the body. Candida is an example of such a dimorphic fungus (Fig. 73-1). It can undergo rapid transformation from the yeast to the hyphal phase in vivo, which partly contributes to its success in invading host tissue.
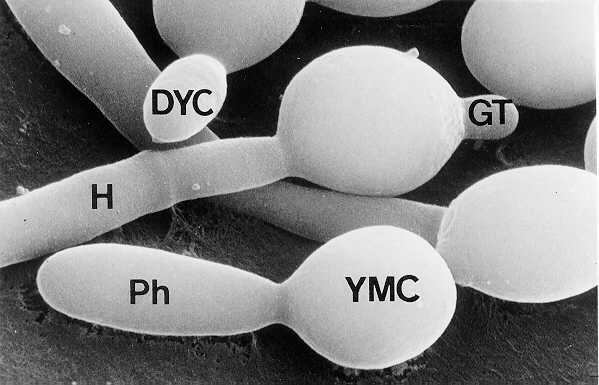
FIGURE 73-1 Dimorphism in C albicans. DYC, Daughter yeast cell; GT, germ tube; H, hypha; Ph, pseudohypha; YMC, yeast mother cell. (X8,980) (From Cole, GT, Kendrick B: Biology of Conidial Fungi. Vol. 1. Academic Press, San Diego, 1981, with permission.)
The true fungi obtain their carbon compounds from nonliving organic substrates (saprophytes) or living organic material (parasites) by absorption of nutrients through their cell wall. Small molecules (e.g., simple sugars and amino acids) accumulate in a watery film surrounding the hyphae or yeast and simply diffuse through the cell wall. Macromolecules and insoluble polymers (e.g., proteins, glycogen, starch, and cellulose), on the other hand, must undergo preliminary digestion before they can be absorbed by the fungal cell. This process involves release of specific proteolytic, glycolytic, or lipolytic enzymes from the hypha or yeast, extracellular breakdown of the substrate(s), and diffusion of the products of digestion through the fungal cell envelope (Fig. 73-2). Fungal pathogens rely on these digestive enzymes to penetrate natural host barriers.
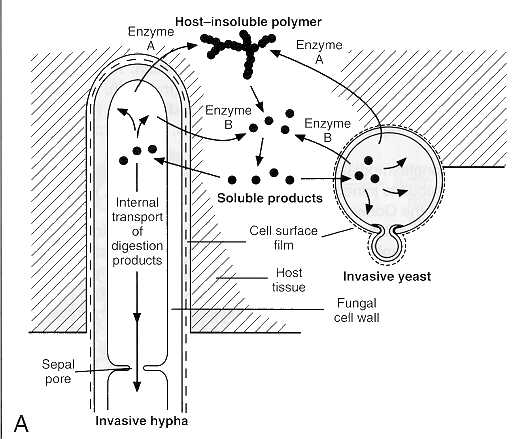
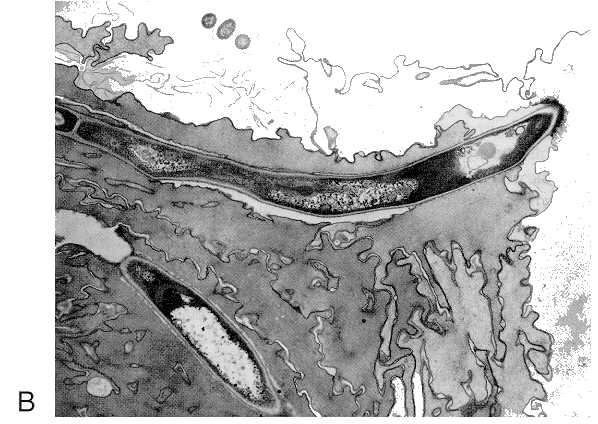
FIGURE 73-2 (A) Extracellular digestion and absorptive nutrition in fungi. (B) Invasive hyphae of C albicans in stratified epithelial tissue of mouse stomach. (X4,250).
Not all species of fungi have cell walls, but in those that do, cell wall synthesis is an important factor in determining the final morphology of fungal elements. Thus, our knowledge of fungal morphogenesis has evolved in parallel with our understanding of fungal cell wall biosynthesis. The fungal wall also protects cells against mechanical injury and blocks the ingress of toxic macromolecules. This filtering effect may be especially important in protecting fungal pathogens against certain fungicidal products of the host. The fungal cell wall is also essential to prevent osmotic lysis. Even a small lesion in the cell wall can result in extrusion of cytoplasm as a result of the internal (turgor) pressure of the protoplast. The composition of fungal cell walls is relatively simple and includes substances not typically found in animal and plant hosts (e.g., chitin). On this basis, it may be possible to identify pathogen-specific molecular targets from investigations of the biosynthesis of fungal wall components. Such targets may prove pivotal for the successful development of antifungal drugs that are not toxic to mammalian cells.
Fungi, like bacteria, are ecologically important as decomposers as well as parasites of plants and animals. Both groups of microbes often inhabit the same ecosystem and thus compete for the same food supply. Associated with this competition is the production by both the fungi and bacteria of secondary products that function as microbial growth inhibitors or toxins. These compounds constitute a rich library of antimicrobial agents, many of which have been developed as pharmacologic antibiotics (e.g., penicillin from Penicillium chrysogenum, nystatin from Streptomyces noursei, amphotericin B from S niveus).
The superficial morphologic similarities between actinomycetes (filamentous bacteria) and molds suggest that the two groups have undergone parallel evolution. Despite the production of branching filaments and mold-like spores, the actinomycetes are clearly prokaryotes, whereas fungi are eukaryotes. Moreover, the sexual reproduction of bacteria, which typically occurs by transverse binary fission, should not be confused with asexual processes of budding and fragmentation associated with mitotic nuclear division in fungi. Most of the molds that produce septate vegetative hyphae reproduce exclusively by asexual means, giving rise to airborne propagules called conidia. On the other hand, elaborate mechanisms of sexual reproduction are also demonstrated by members of the Eumycota. Four distinct kinds of meiospores (products of karyogamy-meiosis-cytokinesis) are recognized: oospores (Oomycetes), zygospores (Zygomycetes), ascospores (Ascomycetes), and basidiospores (Basidiomycetes).
A summary of these and other diagnostic features of the fungi is presented in Table 73-1.

Hyphal growth occurs by extension at the tips. This polarization is at least partially determined by directional movement and accumulation of vesicles that carry wall precursors and wall synthetases to the site of exocytosis at the apical dome of the hypha (Fig. 73-3). Despite the apparent simplicity of hyphal morphogenesis, ultrastructural investigations have shown a sophisticated organization of tip-growth-related organelles and cytoskeletal elements. There is evidence that intussusception and polymerization of chitin microfibrils occur at the apical dome of the hypha and that the biosynthesis of this major cell wall product is controlled by the activity of membrane-bound chitin synthetase. The zymogen form of chitin synthetase has been detected in microvesicles called chitosomes, which appear to transport this enzyme to the hyphal tip. The chitosomes may arise from Golgi-like bodies or by a process of self-assembly of subunits freely within the cytoplasm or within larger vesicular bodies. Activation of chitin synthetase occurs upon fusion of the chitosome with the plasmalemma and may be due to the interaction of a membrane-bound protease and the zymogen. Chitin microfibrillogenesis is initiated at these sites of fusion.
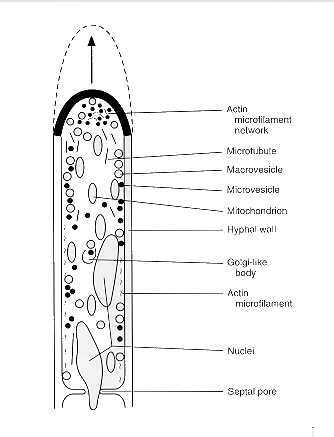
FIGURE 73-3 Polarized growth of hypha.
Evidence has also been presented, primarily from studies of the yeast Saccharomyces cerevisiae, that biosynthesis of skeletal polysaccharides is catalyzed by polysaccharide synthetases (e.g., chitin synthetase and b1-3-glucan synthetase), which are uniformly distributed within the plasmalemma. These wall-synthesizing, cell - membrane-bound enzymes occur in either zymogen or active forms. The model of yeast morphogenesis (Fig. 73-4) suggests that the synthetase is active at sites where the wall is growing and inactive where it is quiescent. One possibility is that microvesicles transport activating factors (e.g., proteases, ATP, and GTP) to the plasmalemma at specific sites of wall biosynthesis (zones of bud emergence and of septal formation). These two concepts of regulation of wall biosynthesis in fungal hyphae and yeasts have been supported by considerable bodies of evidence, and it is likely that both are correct.
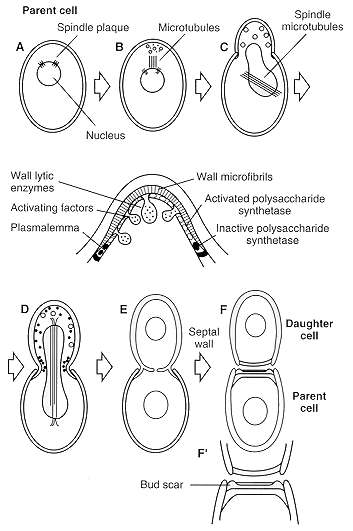
FIGURE 73-4 Stages (A to F) of bud emergence and yeast cell cycle.
Extension growth of hyphal tips and yeast buds logically requires a balance between processes of insertion of newly synthesized polymeric material and modification of the existing microfibrillar matrix to accommodate expansion and further intussusception of wall polymers. In other words, a balance between wall synthesis and wall lysis, or plasticization, is essential for maintaining the orderly processes of hyphal tip elongation and bud emergence. The presence of lytic enzymes in the fungal wall has been reported, including, b1-3-glucanase, N-acetyl-b-D-glucosaminadase, and chitinase. Localization of such activity may be mediated by macrovesicles. These organelles, like microvesicles, are probably derived from Golg-like bodies and are directed to the hyphal tip or yeast bud and fuse with the plasmalemma, thereby delivering their contents to the site of wall synthesis.
Sexual reproduction in the fungi typically involves fusion of two haploid nuclei (karyogamy), followed by meiotic division of the resulting diploid nucleus (Fig. 73-5A). In some cases, sexual spores are produced only by fusion of two nuclei of different mating types, which necessitates prior conjugation of different thalli. This condition of sexual reproduction is known as heterothallism, and the nuclear fusion is referred to as heterokaryosis. Normally plasmogamy (union of two hyphal protoplasts which brings the nuclei close together in the same cell) is followed almost immediately by karyogamy. In certain members of the Basidiomycotina, however, these two processes are separated in time and space, with plasmogamy resulting in a pair of nuclei (dikaryon) contained within a single cell. Karyogamy may be delayed until considerably later in the life history of the fungus. Meanwhile, growth and cell division of the binucleate cell occur. The development of a dikaryotic mycelium results from simultaneous division of the two closely associated nuclei and separation of the sister nuclei into two daughter cells (Fig. 73-5B). An alternative mechanism of sexual reproduction in the fungi is homothallism, in which a nucleus within the same thallus can fuse with another nucleus of that thallus (i.e., homokaryosis). An understanding of these nuclear cycles is fundamental to investigations of fungal genetics.
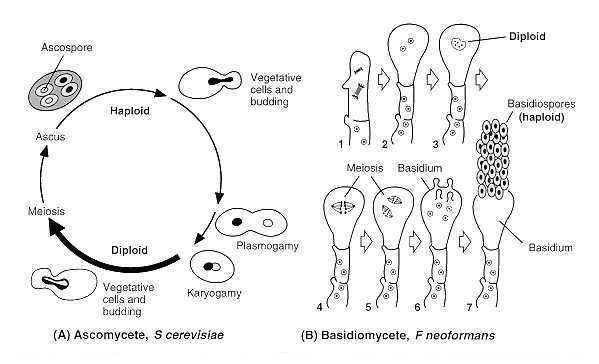
FIGURE 73-5 (A) Life cycle of S cerevisiae. (B) Basidiospore formation by Filobasidiella neoformans, sexual state of Cryptococcus neoformans. (1 and 2) Dikaryon formation. (3) Nuclear fusion (Karyogamy). (4 and 5) Meiosis. (6) Basidiospore formation. (7) Mitosis and basidiospore proliferation.
As mentioned above some fungi are classified as strictly asexually reproducing forms. These include the large group of asexual (imperfect) yeasts (e.g., Candida species) and conidial fungi (e.g., Coccidioides immitis). Most members of this group have permanently lost their ability to produce meiospores. A few undergo rare sexual reproduction, and perhaps for some species we have yet to discover their sexual (perfect) stage. The most common methods of asexual reproduction, in addition to simple budding in yeasts, are blastic development of conidia from specialized hyphae (conidiogenous cells), fragmentation of hyphae into conidia, and conversion of hyphal elements into conidia or chlamydospores (thick-walled resting spores) (Fig. 73-6).
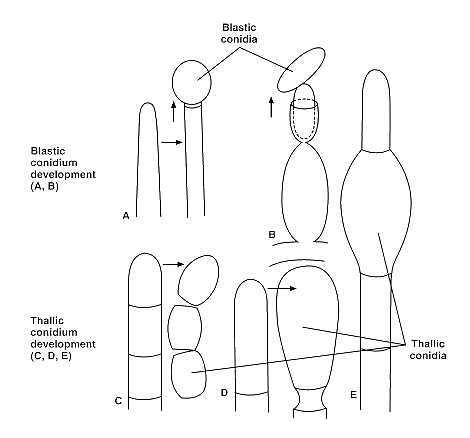
FIGURE 73-6 Methods of asexual reproduction in the conidial fungi. (A) Terminal blastic conidium. (B) Repetitive blastic conidium formation from specialized conidiogenous cell (phialide). (C) Conidium formation by hyphal fragmentation. (D) Conidium formation by conversion of apical segment of hypha into single, asexual propagule. (E) Conversion of hyphal element into an intercalary chlamydospore.
Despite the absence of meiosis during the life cycle of these imperfect fungi, recombination of hereditary properties and genetic variation still occur by a mechanism called parasexuality. The major events of this process (Fig. 73-7) include the production of diploid nuclei in a heterokaryotic, haploid mycelium that results from plasmogamy and karyogamy; multiplication of the diploid along with haploid nuclei in the heterokaryotic mycelium; sorting out of a diploid homokaryon; segregation and recombination by crossing over at mitosis; and haploidization of the diploid nuclei. Sexual and parasexual cycles are not mutually exclusive. Some fungi that reproduce sexually also exhibit parasexuality.
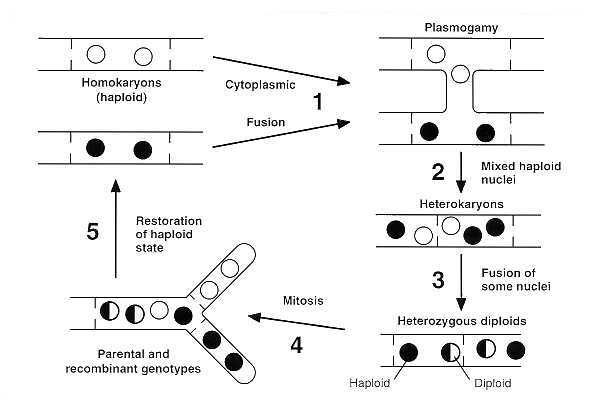
FIGURE 73-7 The parasexual cycle (genetic recombination without meiosis). Stages of the parasexual cycle are numbered as follows (1) Hyphal conjugation (plasmogamy). (2) Heterokaryosis. (3) Nuclear fusion (karyogamy). (4) Mitotic recombination and nondisjunction. (5) Haploidization and nuclear segregation leading to homokaryosis.
An extensive foundation of knowledge on the basic biology of fungi is at hand, including fungi that cause superficial, deep-seated, and systemic infections of humans and other animals. Much less is known, however, of the intricacies of interactions between these largely opportunistic pathogens and their hosts. Many areas of research in medical mycology are still in their infancy and offer formidable challenges and potential rewards. The current application of methods of recombinant DNA technology to problems of fungus-host interactions, especially the identification of pathogenicity genes, holds promise for significant contributions to our knowledge of medically important fungi.
Bulawa CE: Genetics and molecular biology of chitin synthesis in fungi. Ann Rev Microbiol 47:505, 1993
Cole GT, Hoch HC (eds): The Fungal Spore and Disease Initiation in Plants and Animals. Plenum, New York, 1991
Gow NAR, Gadd GM (eds): The Growing Fungus. Chapman and Hall, London, 1995
Kwon-Chung KJ, Bennett JE: Medical Mycology. Lea and Febiger, Philadelphia, 1992
Latgé JP, Boucias D (eds): Fungal Cell Wall and Immune Response. Springer-Verlag, New York, 1991
Odds FC: Candida and Candidosis, 2nd edition. Baillière Tindall, Philadelphia, 1988